Academic Background
William E. Royer, Jr. received his BS from Pennsylvania State University in 1976 and his PhD in biophysics from The Johns Hopkins University in 1984. He was a postdoctoral fellow in Dr. Wayne Hendrickson's laboratory at ColumbiaUniversity and became an associate of the Howard Hughes Medical Institute in 1986 and an associate research scientist at Columbia University in1988. He joined the faculty of the University of Massachusetts MedicalSchool in 1990. He was an established Investigator of the American Heart Association from 1994-1999.

Research Interests
Our laboratory explores the structural basis by which intermolecular interactions regulate biological function in a number of systems. One focus is how assembly of protein subunits regulates their function. Two systems in this area of current interest are interferon regulatory factors, in which phosphorylation triggers dimeric assembly and nuclear translocation, and a dimeric hemoglobin, for which assembly results in substantial cooperative ligand binding. A second focus is the structure-based development of inhibitors for a cancer target, CtBP. These three research areas are described below.
Structural regulation of interferon regulatory factors (IRFs) in the innate immune response
IRF family members play important roles in innate immunity, inflammation and apoptosis. Their clinical importance is evidenced by a strong link between IRF5 and autoimmune diseases, particularly lupus. Our goal is to obtain a structural understanding of activation that will permit insight into approaches for inactivation that could inform development of future therapies in autoimmune disease. The first step in IRF activation is triggered by phosphorylation of Ser/Thr residues in a C-terminal autoinhibitory region. Phosphylation stimulates dimerization, translocation into the nucleus and assembly with the coactivator CBP/p300 and other proteins to stimulate transcription of type I interferons and other target genes. In collaboration with Dr. Celia Schiffer (BMP) and Dr. Kate Fitzgerald (Medicine), we are continuing work on the structural basis for activation of IRFs that was pioneered by our extraordinary colleague, Dr. Kai Lin, prior his tragic death. Our crystal structure of dimeric pseudophosphorylated IRF-5, in comparison with structures of monomeric IRF-3 determined by Dr. Lin, has revealed how phosphorylation triggers a striking conformational rearrangement of the C-terminal region converting it from an autoinhibitory to a dimerization role. Activated dimers are then translocated into the nucleus, where they assemble with transcriptional coactivators to activate transcription.

Figure 1 legend: The crystal structure of the IRF-5 dimer (PDB id code 3DSH) is shown as a ribbon diagram with one subunit in green and one in blue, except for the C-terminal autoinhibition/dimerization region, which is shown in purple for both subunits. Likely phosphorylation sites are shown as small yellow spheres at their alpha-carbon positions. Comparison of the IRF-5 structure with that of monomeric autoinhibited IRF-3 strongly suggests a common mechanism for activation of the interfereon regulatory factors in which phosphorylation triggers a dramatic structural transition of the C-terminal region, resulting in IRF dimerization and exposure of the CBP binding site. These two effects are key steps leading to the transcriptional activation of type 1 interferons and other target genes (see Chen et al. (2008) NSMB 15, 1213-1220).
Structure-based development of inhibitors against CtBP, a cancer target
The paralogous transcription coregulators C-terminal Binding Proteins (CtBP) 1 and 2 are critical modulators of numerous cellular processes. CtBP 1 and 2 have been shown to be overexpressed in many human cancers where they act to inhibit apoptosis and promote metastasis. Unusual among transcription factors, CtBP harbors an essential D-isomer specific 2-hydroxyacid dehydrogenase (D2-HDH) catalytic domain that enables CtBP to be a redox sensor and provides an attractive target for small molecule inhibition. High (mM) concentrations of the CtBP substrate MTOB have been shown by our collaborator, Dr. Steven Grossman at Virginia Commonwealth University, to be able to inhibit tumor growth in cell culture and mouse studies providing proof of principle that CtBP could be an excellent cancer target. By determining the crystal structures of CtBP 1 and 2 in complex with MTOB, we elucidated unique active site features and then used these to identify compounds with substantially higher affinity. Among these features is a water filled hydrophilic cavity, shown below, that is not shared by other D2-HDH family members. We are working to exploit such structural features to design potentially therapeutically useful inhibitors that bind specifically to CtBP with high affinity.
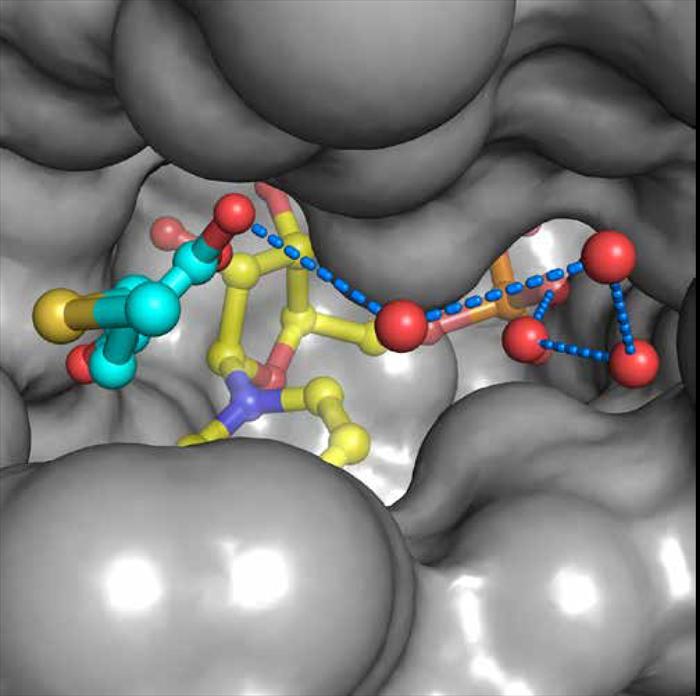
Figure 2 legend: The active site hydrophilic cavity in CtBP1, with the protein cavity surface shown in gray. Four crystallographically observed water molecules, W1-W4, link substrate MTOB (cyan carbon atoms) with an NAD+ (yellow) phosphate. This cavity is not present in other members of the D2-HDH family. Therefore, creating inhibitors with functional groups that bind in this cavity should both gain affinity and specificity for CtBP. (See Hilbert et al. (2014) FEBS Lett. 588, 1743-1748)
Time-resolved crystallographic analysis of cooperative ligand binding
Invertebrate hemoglobins provide a number of useful model systems for exploring how protein subunits communicate. We have investigated systems ranging from the simplest possible allosteric systems to much more complex protein assemblages. As part of our efforts, we determined crystal structures of respiratory proteins with molecular masses of 3.6 x 106 Da comprising 180 different subunits. (Our crystal structure of Lumbricus erythrocruorin was highlighted as the March, 2013 molecule of the month at the protein data bank – see http://www.rcsb.org/pdb/101/motm.do?momID=159.) Our current efforts are focused on applying sub-ns time-resolved crystallographic analysis to a dimeric hemoglobin (Scapharca HbI) to dissect the time-dependent ligand-linked structural changes that underlie cooperative ligand binding. These experiments are being carried out at BioCARS (Advanced Photon Source) in collaboration with Dr. Vukica Srajer and Dr. Zhong Ren. They have already led to a new understanding of the role of water molecules in intersubunit communication as well as permitting the development of a novel dynamic model that incorporates a direct linkage between two active sites. In collaboration with Dr. Francesca Massi (BMP), we are combining the crystallographic results with NMR experiments that investigate the role of interface dynamics in cooperative behavior.